Грибы размножаются вегетативным, бесполым и
половым путями.
Вегетативное
размножение происходит без образования специализированных
органов – любая часть мицелия дает начало новому организму.
Вегетативное размножение применяют при поддержании культуры на
искусственных питательных средах.
Одной из форм вегетативного размножения является образование
хламидоспор – клеток, покрытых толстой оболочкой и способных
переносить длительные периоды голодания, высыхания и другие
неблагоприятные условия среды.
Вегетативным размножением является также почкование клеток и
фрагментация гиф на цилиндрические или овальные клетки –
оидии, образованию которых предшествует появление частых перегородок в
гифах. В благоприятных условиях оидии прорастают в новый мицелий.
Бесполое
размножение грибов происходит при помощи особых спор,
появлению которых не предшествует предварительное слияние клеток и
объединение ядер. У одних грибов споры развиваются эндогенно в
специальных вместилищах, спорангиях, и называются спорангиоспорами. У
других – споры формируются экзогенно на специализированных
гифах мицелия, конидиеносцах, и называются конидиями. Для зигомицетов
характерно бесполое размножение при помощи спорангиоспор; для
аскомицетов, базидиомицетов, дейтеромицетов и некоторых низших грибов
– при помощи конидий.
В отличие от спорангиоспор и спорангиеносцев конидии и конидиеносцы
разнообразны по строению. Конидиеносцы различаются по типу ветвления
(мутовчатое, моноподиальное, дихотомическое, симподиальное), имеют
различные вздутия, стеригмы, фиалиды. Конидиеносцы многих грибов бывают
объединены в коремии, ложа, и пикниды.
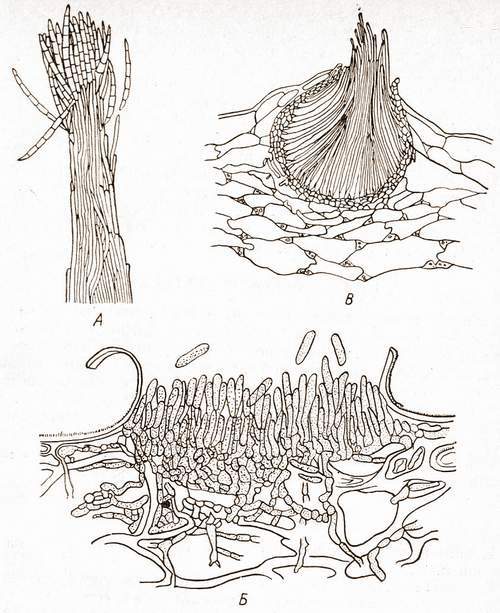
Коремии
представляют собой пучок тесно сближенных конидиеносцев (рис.1).
Подобные образования характерны для некоторых пенициллов. У
дейтеромицетов конидиеносцы часто располагаются тесным слоем на
специальной поверхности, состоящей из плоского сплетения гиф, образуя
ложе. Наиболее сложной формой агрегации конидиеносцев являются
полузамкнутые структуры – пикниды. Они содержат большое
количество конидиеносцев, хорошо защищенных оболочкой.
Рис.1. Конидиеносцы
грибов:
А – коремия Sphaerostilbe;
Б – ложе Gloeosporium;
В – пикнида Septoria
Конидии грибов так же, как конидиеносцы, очень разнообразны по своему
строению. Они бывают одноклеточные и с перегородками; различаются по
форме (шаровидные, эллипсовидные, звездчатые, спирально закрученные и
др.) и цвету (темные, буровато-коричневые, светлые).
Грибы образуют очень большое количество спор бесполого размножения.
Известно, например, что трутовый гриб Ganoderma applanatum
в течение 6 месяцев способен ежедневно высвобождать до 30х106
спор; а
колония пеницилла диаметром 2,5 см может образовать 400х106
конидий.
Распространение спор грибов, вызывающих повреждения материалов, обычно
происходит при помощи ветра, дождя, насекомых, животных и человека.
Половое
размножение установлено для всех групп грибов, кроме
дейтеромицетов, у которых половые стадии отсутствуют. Оно происходит
при помощи специальных спор. Образованию их обязательно предшествует
слияние клеток, часто специализированных, и последующее объединение
ядер.
Половой процесс состоит из трех стадий: плазмогамии, кариогамии и
мейоза. В первую фазу происходит объединение протоплазменного
содержимого двух клеток. Во вторую – слияние ядер и
образование диплоидного ядра с двойным набором хромосом. Третья стадия
характеризуется редукционным делением ядра с восстановлением
гаплоидного набора хромосом. Конечным результатом полового
воспроизведения у всех грибов является образование специальных спор,
служащих для размножения. У низших грибов это споры зародышевого
спорангия, у высших – аскоспоры и базидиоспоры.
Споры, возникшие половым путем, обеспечивают главным образом сохранение
вида в течение периода, неблагоприятного для вегетации, а также
способствуют генетическому разнообразию популяции. Споры бесполого
размножения служат для быстрой колонизации субстрата. Заражение
промышленных материалов происходит в основном с помощью спор бесполого
размножения.
Парасексуальный
процесс. Некоторые грибы не имеют истинного полового
процесса. Обмен генетической информацией у них происходит в результате
парасексуального цикла. Он обнаружен у многих дейтеромицетов и
базидиомицетов главным образом в лабораторных условиях. В результате
этого процесса плазмогамия, кариогамия и гаплоидизация хромосом
происходят не в специализированных половых структурах, или в
определенной стадии жизненного цикла, а в вегетативной гифе мицелия.
Парасексуальный цикл заключается в слиянии генетически различных ядер в
любой вегетативной гифе гетерокариотического мицелия и последующей
рекомбинации участков хромосом при митозе.
Парасексуальный цикл имеет большое значение у несовершенных грибов, для
которых характерно отсутствие полового процесса. Например, для грибов
рода Verticillium
он является источником возникновения изменчивости. У
некоторых грибов парасексуальный цикл происходит и при наличии полового
процесса.